Кора больших полушарий головного мозга
Кора больших полушарий головного мозга представляет собой слой серого вещества толщиной 1—5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга, развившаяся на поздних этапах эволюции животного мира, играет исключительно важную роль в осуществлении психической, или высшей нервной деятельности, хотя эта деятельность является результатом работы мозга как единого целого. Благодаря двусторонним связям с нижележащими отделами нервной системы, кора может участвовать в регуляции и координации всех функций организма. У человека кора составляет в среднем 44% от объёма всего полушария в целом. Её поверхность достигает 1468—1670 см2.
Строение коры. Характерной особенностью строения коры является ориентированное, горизонтально-вертикальное распределение составляющих её нервных клеток по слоям и колонкам; таким образом, корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними. Пространство между телами и отростками нервных клеток коры заполнено нейроглией и сосудистой сетью (капиллярами). Нейроны коры подразделяются на 3 основных типа: пирамидные (80—90% всех клеток коры), звездчатые и веретенообразные. Основные функциональный элемент коры — афферентно-эфферентный (т. е. воспринимающий центростремительные и посылающий центробежные стимулы) длинноаксонный пирамидный нейрон . Звездчатые клетки отличаются слабым развитием дендритов и мощным развитием аксонов, которые не выходят за пределы поперечника коры и охватывают своими разветвлениями группы пирамидных клеток. Звездчатые клетки выполняют роль воспринимающих и синхронизирующих элементов, способных координировать (одновременно тормозить или возбуждать) пространственно близкие группы пирамидных нейронов. Корковый нейрон характеризуется сложным субмикроскопическим строением. Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Все эти показатели определяют архитектуру коры, или её цитоархитектонику.
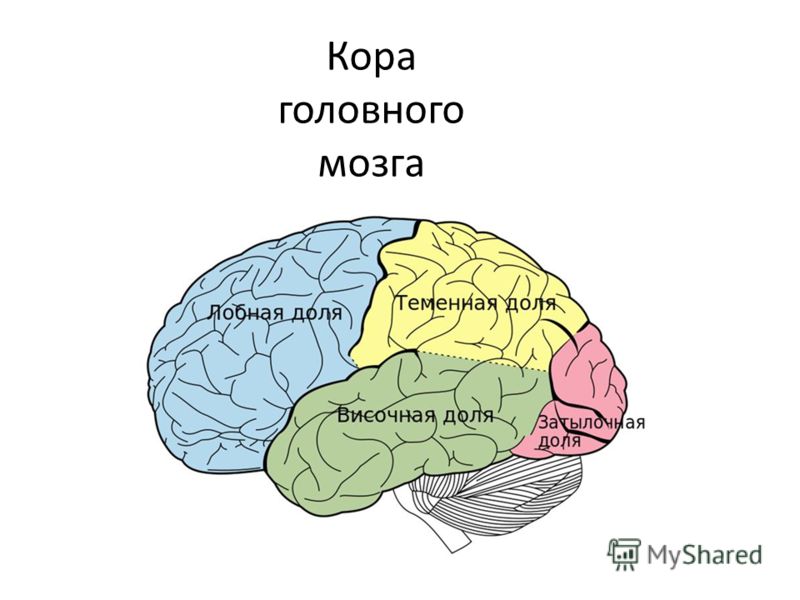
Наиболее крупные подразделения территории коры — древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и межуточная кора. Поверхность новой коры у человека занимает 95,6%, старой 2,2%, древней 0,6%, межуточной 1,6%.
Если представить себе кору мозга в виде единого покрова (плаща), одевающего поверхность полушарий, то основная центральная часть его составит новая кора, в то время как древняя, старая и межуточная займут место на периферии, т. е. по краям этого плаща. Древняя кора у человека и высших млекопитающих состоит из одного клеточного слоя, нечетко отделённого от нижележащих подкорковых ядер; старая кора полностью отделена от последних и представлена 2—3 слоями; новая кора состоит, как правило, из 6—7 слоев клеток; межуточные формации — переходные структуры между полями старой и новой коры, а также древней и новой коры — из 4—5 слоев клеток. Неокортекс подразделяется на следующие области: прецентральную, постцентральную, височную, нижнетеменную, верхнетеменную, височно-теменно-затылочную, затылочную, островковую и лимбическую. В свою очередь, области подразделяются на подобласти и поля. Основной тип прямых и обратных связей новой коры — вертикальные пучки волокон, приносящие информацию из подкорковых структур к коре и посылающие её от коры в эти же подкорковые образования. Наряду с вертикальными связями имеются внутрикортикальные — горизонтальные — пучки ассоциативных волокон, проходящие на различных уровнях коры и в белом веществе под корой. Горизонтальные пучки наиболее характерны для I и III слоев коры, а в некоторых полях для V слоя. Горизонтальные пучки обеспечивают обмен информацией как между полями, расположенными на соседних извилинах, так и между отдалёнными участками коры (например, лобной и затылочной).
{kind=link}
Функциональные особенности коры обусловливаются упомянутым выше распределением нервных клеток и их связей по слоям и колонкам. На корковые нейроны возможна конвергенция (схождение) импульсов от различных органов чувств. Согласно современным представлениям, подобная конвергенция разнородных возбуждений — нейрофизиологический механизм интегративной деятельности головного мозга, т. е. анализа и синтеза ответной деятельности организма. Существенное значение имеет и то, что нейроны сведены в комплексы, по-видимому, реализующие результаты конвергенции возбуждений на отдельные нейроны. Одна из основных морфо-функциональных единиц коры — комплекс, называемый колонкой клеток, который проходит через все корковые слои и состоит из клеток, расположенных на одном перпендикуляре к поверхности коры. Клетки в колонке тесно связаны между собой и получают общую афферентную веточку из подкорки. Каждая колонка клеток отвечает за восприятие преимущественно одного вида чувствительности. Например, если в корковом конце кожного анализатора одна из колонок реагирует на прикосновение к коже, то другая — на движение конечности в суставе. В зрительном анализаторе функции восприятия зрительных образов также распределены по колонкам. Например, одна из колонок воспринимает движение предмета в горизонтальной плоскости, соседняя — в вертикальной и т. п.
Второй комплекс клеток новой коры — слой — ориентирован в горизонтальной плоскости. Полагают, что мелкоклеточные слои II и IV состоят в основном из воспринимающих элементов и являются «входами» в кору. Крупноклеточный слой V — выход из коры в подкорку, а среднеклеточный слой III — ассоциативный, связывающий между собой различные корковые зоны.
Локализация функций в коре головного мозга
Современная концепция о локализации функций в коре большого мозга выходит из учения И.П. Павлова о динамической локализации функций. На основании экспериментальных исследований И.П. Павлов показал, что кора большого мозга представлена совокупностью анализаторов, где каждый из них имеет центральную зону — ядро анализатора и периферическую, где корковое представительство является рассеянным. Вследствие такой структуры анализатора корковые зоны его как бы перекрывают одна другую и образуют тесно связанное морфофункциональное объединение. Динамическая локализация функций в коре предусматривает возможность использования одних и тех же структур мозга для обеспечения разных функций. Это означает, что в выполнении той или другой функции принимают участие разные отделы коры большого мозга. Например, такие высшие психические процессы, как речь, письмо, чтение, счет и т.п., никогда не осуществляются одним изолированным центром, а опираются на сложную систему совместно функционирующих зон головного мозга. Динамическая локализация функций не исключает наличие центров в коре большого мозга, но их функция определяется связями с другими участками коры.
Необходимо отметить, что степень локализованности разных функций коры неодинаковая. Только элементарные корковые функции, которые обеспечиваются отдельными анализаторами, первичными рецепторными аппаратами, можно связать с соответствующими участками коры. Сложные, филогенетически молодые функции не могут быть узко локализованными; в их осуществлении участвуют большие участки коры большого мозга или даже кора в целом.
Дальнейшее развитие учения о динамической локализации функций в коре получило в работах П.К. Анохина (1955), который сформулировал концепцию функциональных систем высших мозговых функций. В соответствии с современными представлениями функциональная система имеет сложное иерархическое строение. Она включает в разных соединениях корковые, подкорковые центры, проводящие пути, исполнительные органы. Причем одни и те же нервные образования могут быть составными разных функциональных систем. Непосредственно та или другая высшая мозговая функция реализуется благодаря сложному, упорядоченному, динамическому взаимодействию разных систем мозга.
Значительный вклад в понимание функциональной организации коры большого мозга внесли исследования канадского нейрохирурга У. Пенфильда (1964), проведенные во время оперативного вмешательства на мозге человека. Основным принципом функциональной организации проекционных систем в коре является принцип топической локализации, которая основывается на четких анатомических связях между отдельными воспринимающими элементами периферии и корковыми клетками проекционных зон. В каждой из этих систем анализаторов в зависимости от отношения разных участков коры к другим образованиям мозга различают три типа корковых полей (Г.И. Поляков, 1973).
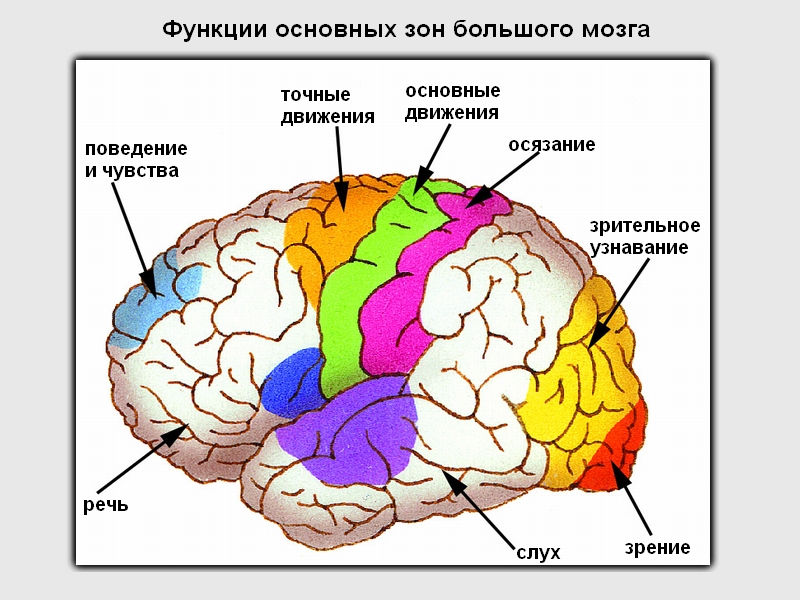
Первичные проекционные поля отвечают тем архитектоническим участкам, в которых локализуются корковые отделы анализаторов: анализатора общей чувствительности — в постцентральной извилине, обонятельного и слухового в височной доле, зрительного в затылочной. С этими полями связаны простые, элементарные функции: общая чувствительность кожи, слух, обоняние, зрение. Это поля, которые не могут обеспечить интегративную функцию восприятия, они лишь реагируют на определенные раздражения одной модальности и не отвечают на раздражение другой. В первичных проекционных полях самыми развитыми являются нейроны IV афферентного слоя. Для первичных проекционных полей характерен соматотопический принцип строения, т. е. представительство чувствительных функций в определенных зонах коры.
{kind=link}
Вторичные проекционные поля расположены вокруг первичных. Они непосредственно не связаны со специфическими проводящими путями. Во вторичных корковых полях преобладают нейроны второго и третьего слоев коры; здесь имеется большое количество мультисенсорных нейронов, которые обеспечивают, по сравнению с первичными полями, другой характер реагирования. Электрическое раздражение вторичных проекционных полей вызывает у человека сложные зрительные образы, мелодии, в отличие от элементарных ощущений (вспышка, звук), которые возникают в случае раздражения первичных полей. Во вторичных проекционных полях происходит высший анализ и синтез, более подробная обработка информации, осознание ее.
Вторичные проекционные поля вместе с первичными составляют центральную часть анализатора, или его ядро. Взаимодействие нейронов этих зон носит сложный, неоднозначный характер, и в условиях нормальной деятельности мозга оно основывается на последовательном изменении возбудительных и тормозных процессов в соответствии с характером конечного результата. Это и обеспечивает динамические свойства локализации.
Описанная функциональная организация коры в виде четко разделенных по принципу модальной специфичности полей в наибольшей мере выражена у человека и высших представителей животного мира. В частности, у человека вторичные проекционные поля составляют около 50 % всей коры большого мозга (у обезьян — около 20 %).
Третичные проекционные поля — это ассоциативные зоны, которые размещены в местах перекрывания отдельных анализаторов. Различают две основных ассоциативных зоны: в лобной доле перед прецентральной извилиной и на границе между вторичными проекционными полями теменной, затылочной и височной долей.
Третичные проекционные поля, или зоны перекрытия, не связаны непосредственно с периферическими рецепторными аппаратами, но они тесно связаны с другими участками коры, в том числе и с проекционными полями. Сюда поступают также сигналы от ассоциативных ядер таламуса.
Третичные проекционные поля имеют непосредственное отношение к высшим психическим функциям. С функцией этих зон с%D